Leuchtende Karnivoren: Die Lumineszenz der Schierlingsbecher Siegfried R. H. Hartmeyer1, Richard Bayerl2 und Irmgard Hartmeyer1 1)DE-79576 Weil am Rhein. 2) DE-70176 Stuttgart. Publiziert in Das Taublatt Heft 75:33-44 (2013) Schlüsselworte: Fluoreszenz, Lumineszenz, Sarracenia, Nepenthes, Coniin, Karnivoren Vorgeschichte Ende 2012 erschien in der Zeitschrift Plant Biology ein Artikel mit dem Titel Fluorescent prey traps in carnivorous plants
(Kurup et al. 2012), was auf DeutschFluoreszierende Beutefallen bei Karnivoren
bedeutet. Bei BBC Nature News wurde
daraus im Februar 2013 bereits: Carnivorous plant species glow blue to lure
prey (Michelle Warwicker
2013), auf Deutsch: Karnivorenarten
leuchten blau um Beute anzulocken. Die indischen Autoren schlussfolgern aus
ihren Versuchen an Nepentheskannen mit gewaschenen/abgeschnittenen Peristomen
verglichen mit naturbelassenen Peristomen sogar, dass diese Lumineszenz den
Fangerfolg der Pflanzen deutlich steigert. Donnerwetter sagten sich da auf
Anhieb diverse Karnivorenfreunde und richteten im Dunkeln begeistert
Geldscheinprüfer und andere Schwarzlichtlampen auf alles, was
Blattfallen hat,
um das faszinierend bläuliche Schimmern der Fallen mit eigenen
Augen zu sehen. Als Beleg für die Wahrhaftigkeit des
Phänomens stellten sie anschließend an den
Film Avatar erinnernde Fotos ins Internet. Aber Vorsicht, ganz so
simpel ist
die Geschichte dann doch nicht. Deshalb griffen die Autoren (bei
Richard und
Siggi gehörte UV-Spektroskopie zur Ausbildung als Chemielaborant)
selbst zu
einer UV-Leuchte mit definierten Wellenlängen und dokumentierten
im Gewächshaus
der Hartmeyers das kalte Leuchten der Fallen. Um das Geschehen auch
für
Nichtfachleute besser verständlich zu machen, beschreiben wir erst
einmal die
wichtigsten bekannten Leuchterscheinungen anhand von Beispielen. Wer
sich
detailliert über die physikalischen Hintergründe der
Quantensprünge von
Elektronen informieren möchte, die hier den Rahmen des Artikels
sprengen
würden, braucht dazu nur die Schlagwörter am Anfang des
nächsten Abschnitts zu
googeln.
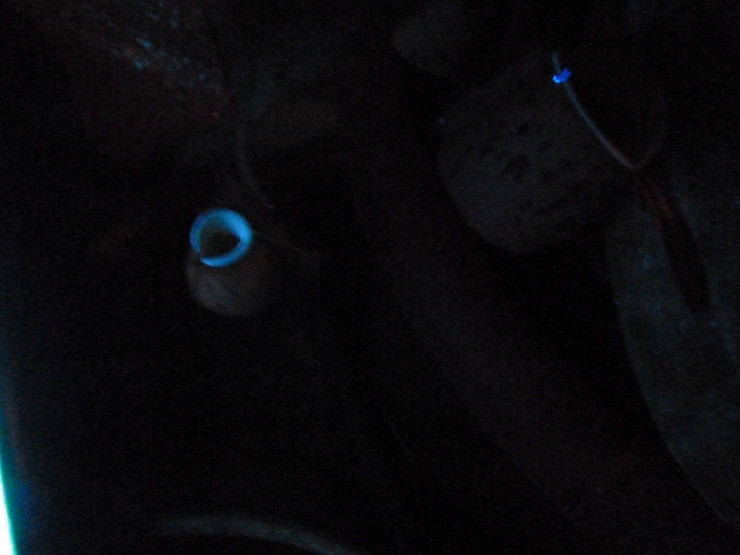
Einige wichtige Leuchterscheinungen Die Lumineszenz verschiedener Materialien wird auch als „kaltes Licht“ bezeichnet. Sie benötigt eine Anregungsenergie, die aus sehr unterschiedlichen Energieformen bestehen kann, welche daraufhin als elektromagnetische Strahlung in Form von Licht auch bei niedrigen Temperaturen wieder emittiert wird. Die Art der Anregungsenergie bestimmt dabei die genaue Bezeichnung der unterschiedlichen Lumineszenzen. Das Glühen, also die Lichtemission heißer Körper mit Temperaturen ab etwa 600 °C (rotglühend), wird nicht als Lumineszenz bezeichnet. Die Lichtfarbe wird jedoch in allen Fällen in Grad Kelvin gemessen (Sonnenlicht = 5500 °K). Karnivorenfreunde, die derzeit mit neuartigen LEDs als Kunstlicht für ihre Pflanzen experimentieren, kennen diese Kelvinskala meist recht gut, da eine optimale Lichtfarbe für ein gesundes Gedeihen äußerst wichtig ist. Die Elektrolumineszenz bringt die oben erwähnten sparsamen LEDs zum Leuchten, aber auch Strom fressende Metalldampflampen. Dabei werden Stoffe durch das Anlegen einer elektrischen Spannung dazu veranlasst, Licht abzustrahlen (siehe auch Piezolumineszenz unten). Diese Beleuchtungsform ist energieeffizient, besonders im Vergleich mit herkömmlichen Glühlampen, die bis zu 97 % des Stroms als Wärme abstrahlen. LED-Lampen sind zwar noch teuer, aber ungiftig im Gebrauch, weshalb sie den sogenannten „Energiesparlampen“ mit Leuchtröhrenprinzip unbedingt vorzuziehen sind, die bei höherem Stromverbrauch auch noch erhebliche Mengen an hochgiftigem Quecksiber enthalten. Tatsächlich können bereits beim Zerbrechen einer einzigen „Energiesparlampe“ in ungenügend gelüfteten Räumen äußerst gesundheitsschädliche Konzentrationen des Nervengifts eingeatmet werden. Die Piezolumineszenz (auch Quarzleuchten) ist eine Besonderheit der Elektrolumineszenz und soll in diesem Zusammenhang noch erwähnt werden, weil sie sehr alltäglich ist. Wer immer ein modernes Feuerzeug ohne Feuerstein benutzt, erzeugt den Zündfunken durch sogenannte Piezoelektrizität, die bei der Reibung von Quarzkristallen, aber auch anderen Materialien wie z. B. Bariumtitanat, entsteht. Dieser Piezostrom ist nicht nur die Ursache des Zündfunkens, sondern dient auch als Anregungsenergie für das bei Piezomaterialien unter Druck typische Aufleuchten. Die Phosphoreszenz ist im Prinzip eine Variante der Fluoreszenz (siehe unten), benannt nach dem Leuchten des weißen Phosphors, bei dem es sich jedoch, wie man heute weiß, in Wirklichkeit um eine Chemolumineszenz (siehe unten) handelt. Die Anregungsenergie ist UV-Licht, welches jedoch über einen Zeitraum von Sekunden bis zu mehreren Stunden gespeichert wird, bis die Energie schließlich wieder als sichtbares Licht abgestrahlt wird. Im Handel sind darauf beruhende Leuchtfarben erhältlich, sowie viele im Dunkeln leuchtende Gegenstände, vom Hinweisschild über Leuchtbecher für die Bar, bis zu Leuchtkieseln für den Gartenweg. Meist sind diese aus Kuststoff. Stellt man diese Gegenstände tagsüber ans Licht, leuchten sie anschließend im Dunkeln mehrere Stunden lang grünlich. Das früher fälschlicherweise als Phosphoreszenz gedeutete geheimnisvolle grüne Leuchten an vermodernden Baumstämmen in Mooren ist übrigens, wie man heute weiß, eine Biolumineszenz (siehe unten), verursacht durch das Mycel eines Bäume parasitierenden Pilzes, dem Hallimasch. Die Fluoreszenz funktioniert, wie oben bereits erwähnt, im Prinzip wie die Phosphoreszenz, allerdings wird die Anregungsenergie in Form von UV-Strahlung nicht gespeichert, sondern umgehend in sichtbares Licht umgewandelt und emittiert. Am Bekanntesten ist sicher der Effekt von Fluoreszeinlösung, die gern auf Labortischen in Kinofilmen aus allerlei chemischem Gerät sehr wirkungsvoll gelbgrün hervorleuchtet. Fast täglich haben wir Kontakt mit bläulich fluoreszierenden optischen Aufhellern, die uns Papier oder Textilien als „superweiß“ erscheinen lassen. Diese bläuliche Fluoreszenz werden wir auch bei unseren Lumineszenzexperimenten an Karnivoren antreffen. Das Chlorophyll der Pflanzen fluoresziert intensiv rot und seit dem Großbrand bei Sandoz in Schweizerhalle am 1. November 1986 kennt die Welt auch den Farbstoff Rhodamin B. Als fluoreszierender Markierungsfarbstoff war er hochgiftigen Agrochemikalien zugemischt worden, um kleinste Kontaminationen anzuzeigen. Aus einer brennenden Chemielagerhalle gelangte er mit großen Mengen Löschwasser in den Rhein, wo die Intensität der Fluoreszenz ausreichte, den Rhein auf Luftaufnahmen von Basel über Hunderte Kilometer bis fast nach Holland bei Sonne rot leuchten zu lassen. Bei allen Beispielen genügen bereits sehr kleine Konzentrationen für eine deutlich sichtbare Fluoreszens. Substanzen mit roten, grünen und blauen Lumineszenzen lassen sich auch kombinieren, wodurch sich alle Mischfarben bis hin zu Weiß erzeugen lassen. Die Chemolumineszenz unterscheidet sich von Fluoreszenz und Phosphoreszenz durch die Anregungsenergie, die nicht aus UV-Licht besteht, sondern das Resultat einer chemischen Reaktion ist. Anwendung findet sie beispielsweise in Leuchtstäben (Knicklichter) die drei Chemikalien enthalten: Oxalsäurester, fluoreszierenden Farbstoff und Wasserstoffperoxid. Das Peroxid befindet sich in einer Ampulle, die beim Gebrauch durch Knicken zerbrochen wird, wodurch alle Reagenzien vermischt werden und die Reaktion beginnt. Die Art des Farbstoffs kann variiert werden und bestimmt die Farbe der Lumineszenz, solange die chemische Reaktion die notwendige Anregungsenergie liefert, was mehrere Stunden andauern kann. In der Biochemie wird diese Form der Lumineszenz auch zum Markieren bestimmter Verbindungen genutzt, die daraufhin aufgrund der Empfindlichkeit der Methode bis auf molekularer Ebene nachgewiesen werden können. Die Biolumineszenz ist eine Form der Chemolumineszenz, die in lebenden Organismen auftritt (Bio=Leben, Lumineszenz = Leuchten). Dieses Phänomen kennt man von den Glühwürmchen, von diversen Tiefseetieren, etwa 40 Arten von Pilzen, wie dem oben erwähnten Hallimasch sowie Plankton und Bakterien. Die Radiolumineszenz wird durch Alpha-, Beta-, oder Röntgenstrahlung als Anregungsenergie ausgelöst, wodurch viele Materialien, z. B. Radium mit Zinksulfid, zu leuchten beginnen. Seit Anfang des vorigen Jahrhunderts wurden auf diesem Prinzip beruhende Leuchtfarben hergestellt. Zifferblätter von Uhren wurden damit bemalt - und im Nachhinein besonders bedenklich, auch ganze Leuchtbilder im Postkartenformat. Diese fanden in den 1960er Jahren, in denen die „saubere Atomkraft“ als die Lösung aller Energieprobleme gepriesen wurde, durch ihr geheimnisvolles nächtliches Leuchten besonders in Kinderzimmern großen Anklang (pers. Erfahrung). Im Müll, in dem das alles hoffentlich inzwischen gelandet ist, könnte diese Form der Lumineszenz aufgrund der Halbwertszeit des Radiums (1600 Jahre) theoretisch noch viele Jahrtausende lang weiter leuchten. Die Dokumentation der Lumineszenz Um die Lumineszenz von Karnivoren zu untersuchen, benutzten wir eine moderne LED-Schwarzlicht-Taschenlampe mit einer Wellenlänge von 390 nm und eine echte UV-Leuchte (Hanau Fluotest) mit zwei separat zu schaltenden Wellenlängen im nahen (366 nm) und mittleren (254 nm) UV-Bereich. Die benutzt Richard üblicherweise für seine Untersuchungen an fluoreszierenden Mineralien. Wir warteten ab, bis es abends in den Gewächshäusern dunkel genug war, um die Lumineszenz überhaupt sehen zu können, und leuchteten alle möglichen Karnivoren aus kurzer Entfernung mit den verschiedenen Wellenlängen an. Bereits im Vorjahr hatten wir das mit der LED-Schwarzlichtlampe gemacht und tatsächlich sorgte das dunkelblaue Leuchten, das mit 390 nm für das menschliche Auge genau im Grenzbereich zwischen unsichtbarer UV-Strahlung (10 – 380 nm) und sichtbarem Licht (380 – 780 nm) liegt, umgehend für ein faszinierendes bläuliches Avatar-Ambiente. Allerdings war bei Weitem nicht alles, was da exotisch schimmerte, auch echte Lumineszenz!
Tatsächlich zeigen hohe Konzentrationen fluoreszierender Verbindungen, ob in Mineralien oder auf Geldscheinen (die extra für einen Test im Geldscheinprüfer spezielle optische Aufheller enthalten) eine helle Fluoreszenz bei dieser Anregungswellenlänge. Aber das Spektrum der Diodenleuchte enthält auch einen erheblichen Anteil an sichtbarem blauen Licht (etwa 410 nm), welches von hellen Flächen mehr und von dunklen deutlich weniger reflektiert wird. Durch diese starken Kontraste neigen besonders Laien dazu, die helleren, stärker reflektierenden Flächen als Fluoreszenz zu deuten, obwohl es sich nur um Reflexion handelt. Mit der LED-Leuchte lässt sich zwar rote und grüne Fluoreszenz recht gut erkennen, aber eine bläuliche nur zweifelsfrei, wenn sie intensiv genug ist, um den sichtbaren Anteil des reflektierten Blaus deutlich zu überstrahlen. Zwar gab es einige Stellen im Sphagnummoos, die offensichtlich fluoreszierten, auch diverse Nektarien an Nepenthes sowie die Trichome einiger Sonnentau, aber in den meisten Fällen interpretierten wir das Leuchten eher als Reflexion und sahen daher 2012 keinen besonderen Grund über das Phänomen einen Bericht zu schreiben, zumal die Fluoreszenz bei Pflanzen an sich gar keine seltene Erscheinung ist. 2013 hatte sich die Situation jedoch aufgrund der anfangs genannten Publikation von Kurup et al. (2012) geändert, die im Internet viel diskutiert und sogar im Magazin Der Spiegel (2013) mit Bild erläutert wurde. Daher beschlossen wir, die Versuche mit einer professionellen UV-Leuchte mit kürzerer Wellenlänge zu wiederholen. Die indischen Autoren benutzten für ihre Versuche eine Anregungswellenlänge von 366 nm (Nahes UV), die aufgrund der kürzeren Wellenlänge energiereicher ist und weniger sichtbare Blauanteile enthält, als das LED-Schwarzlicht. Dadurch wird auch die bläuliche Lumineszenz deutlich intensiver und besser sichtbar. Die von uns verwendete UV-Leuchte verfügt über diese 366 nm und ist zusätzlich umschaltbar auf eine noch energiereichere Wellenlänge von 254 nm (Mittleres UV), was jedoch bei unseren Versuchen die Fluoreszenz gegenüber 366 nm nicht weiter intensivierte, sondern mitunter sogar eher etwas schwächer ausfallen ließ. Wir dokumentierten unsere Versuche mit einer Lumix DMC-TC10, bei der die Blitzfunktion natürlich ausgeschaltet wurde. Die beigefügten Fotos zeigen alle die Lumineszenz bei 366 nm Anregungswellenlänge und wurden farblich nicht aufgearbeitet, lediglich aufgrund der Grobkörnigkeit durch Unterbelichtung etwas nachgeschärft. Tatsächlich zeigte sich mit den kürzeren Anregungswellenlängen 2013 bei vielen Nepenthes eine sehr deutliche Lumineszenz im oberen Bereich der Kannen am Peristom, an den Zuckeragglomerationen um die Nektarien der Deckelunterseite, sowie fast punktförmig aber sehr kontrastreich an den über die Kannenoberfläche verteilten Nektarien. Bestätigen können wir ein kaltes Leuchten also eindeutig bei N. alata, N. ampullaria (Bild1), N. albomarginata (Bild 1), bei der übrigens auch das arttypische weiße Band unter dem Peristom hellweiß leuchtet, N. bicalcarata (Bild 3), N. khasiana (Bild1), die auch in der Publikation von Kurup et al. (2012) untersucht wurde, N. mirabilis (Bild 3), N. x mixta (N. northiana x maxima), N. pervillei, N. rafflesiana (Bild 1), N. talangensis x truncata (Bild 1 & 3), N. truncata (Bild 1) und N. x ventrata (N. ventricosa x alata). Nur als sehr kleine Flecken war die Lumineszenz auf dem Peristom von N. attenboroughii sichtbar. N. jacqueliniae (Bild 2), N. gymnamphora (Bild 2) sowie N. veitchii (Bild 2) reagierten negativ.
Was die anderen Gattungen in unserer Sammlung betrifft, zeigten die Blattränder von Brocchinia reducta eine leichte Lumineszenz. Gar kein Leuchten gab es auf den Blattoberflächen unserer Heliamphora minor, verschiedener Utricularia wie beispielsweise U. dichotoma, U. calycifida, U. longifolia, U. reniformis, U. subulata, sowie aller Pinguicula wie P. agnata, P. ehlersiae, P. esseriana, P. gigantea, P. moctezumae, P. x sethos. Völlig negativ waren auch die Klebetröpfchen verschiedenster Sonnentau, um nur einige aufzuzählen: D. capensis, D. capensis x aliciae, D. cistiflora, D.felix, D. glanduligera, D. indica, D. madagascariensis, D. tokaiensis, D. trinervia, diverse rosettige Knollendrosera wie D. bulbosa, D. erythrorhiza, D. lowriei, D. whittakeri, D. tubaestylus und hochwachsende Arten wie D. macrantha, D. modesta, D. peltata, D. stricticaulis. Lediglich die dichte Behaarung der Petiolen von Drosera ordensis fluoreszierte sehr eindeutig. Es ist daher wahrscheinlich, dass dies auch bei anderen Arten der Sektion Lasiocephala der Fall ist, von denen wir jedoch nur wenige Pflanzen besitzen, die obendrein zum Zeitpunkt der Versuche, wie beispielsweise D. falconeri, eingezogen waren. Bei den Zwergsonnentau D. dichrosepala, D. enodes, D. ericksoniae, D. pulchella, D. scorpioides war das Leuchten der Haare im Zentrum nicht ganz eindeutig. Es gibt dazu jedoch eine glaubhafte Aufnahme mit UV-Licht im CPUK-Forum, bei der die Trichome im Zentrum von D. scorpioides recht hell weiß aufzuleuchten scheinen, ganz ähnlich wie die Behaarung von D. ordensis und das weiße Band unter dem Peristom von N. albomarginata. Leider waren im März unsere Aldrovanda, Dionaea und Sarracenia in Winterruhe, weshalb hier eine Untersuchung aktiver Fallen nicht möglich war. Bei Sarracenia ist ein fluoreszierender Wirkstoff im Nektar allerdings schon länger bekannt (siehe Diskussion), daher zeigen sie sicher eine ähnliche Fluoreszenz wie die positiv getesteten Nepenthes und wurden ja auch im Originalartikel dahingehend als positiv beschrieben. Zusammenfassend lässt sich sagen, dass bei vielen, jedoch nicht allen Nepenthesarten, in dunkler Umgebung bei Anregungsenergien von 254 und 366 nm eine deutliche Lumineszenz in den Bereichen sichtbar ist, an denen sich Nektarien mit Zuckeragglomeraten befinden, oder am Peristom, wo sich der in Feuchtigkeit gelöste Zucker der Nektarien durch Benetzung des Gewebes offensichtlich recht gleichmäßig flächig verteilt. Die fluoreszierende Substanz ist also eindeutig ein Bestandteil des Nektars. Außerdem zeigten die Blattränder von B. reducta, einige Behaarungen an Drosera, jedoch nicht deren Tentakel sowie der weiße Ring unter dem Peristom von N. albomarginata eine Lumineszenz, die aber offensichtlich auf anderen Ursachen beruht, als die des Nektars.
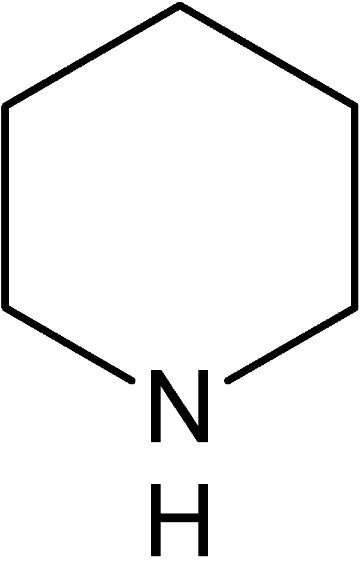
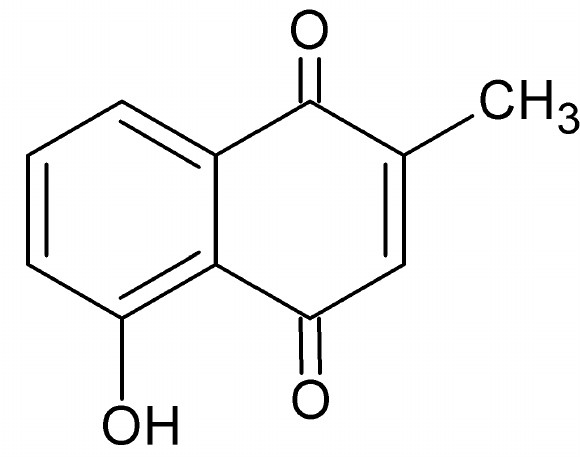
Lumineszenz ist in Fauna und Flora ein weitverbreitetes Phänomen. Isoliert man Chlorophyll, leuchtet es unter UV-Licht rot und Blut fluoresziert intensiv bläulich, wie die Verwendung von Schwarzlicht zur Beweisaufnahme an Tatorten beweist. Nicht nur in industriell hergestellten optischen Aufhellern, auch in der Natur finden sich verbreitet Substanzen, deren Moleküle beispielsweise aus bestimmten Phenolen, polyzyklischen (mehrkernigen) oder heterozyklischen (Kohlenstoff ist im Ring teilweise durch andere Atome ersetzt) Aromaten bestehen, die oft deutlich fluoreszieren. Unsere Experimente bestätigen zwar eindeutig die von Rayani Kurup et al. (2012) beschriebene Fluoreszenz bei diversen Karnivoren, deren Beweisführung für eine messbare Zunahme des Beutefangs aufgrund der nur im Dunkeln sichtbaren Lumineszenz muss jedoch mit Skepsis betrachtet werden. Bei Tageslicht wird das kalte Leuchten auf farbigem Untergrund schlicht überstrahlt, lediglich weiße Strukturen könnten durch bläuliche Fluoreszenz unter Umständen etwas intensiver weiß wirken. Ein Effekt, der für Papier und Textilien durch den Einsatz optischer Aufheller genutzt wird, um das Weiß der behandelten Materialien als „Superweiß“ erscheinen zu lassen. Die Industrie benutzt dafür oft Stilben-Triazon-Verbindungen, die je nach Hersteller als Leucophor™ oder Blankophor™ (Stilben Aufheller) im Handel sind. Ob eine sichtbare Lumineszenz durch die geringen UV-Anteile des Mondlichts ausgelöst wird, die in der Lage wäre, nachtaktive Arthropoden anzulocken, erscheint uns recht unwahrscheinlich. Feldversuche dazu fehlen. Da die Lumineszenz jedoch offenkundig nicht vom Zellgewebe ausgeht, sondern vom Nektar, der durch seinen Zuckergehalt völlig unstrittig bereits ein äußerst effektives Anlockungsmittel bei fast allen Blütenpflanzen ist, muss dessen Fluoreszenz wohl eher als Nebeneffekt einer optisch aktiven Substanz gesehen werden, die möglicherweise einen ganz anderen, nämlich einen pharmakologischen Effekt auf Beutetiere hat. Die experimentelle Beweisführung der Autoren bezüglich des erhöhten Beutefangs bei Fallen mit Lumineszenz bestätigt im Grunde nur die Wirkung des Nektars. Dieser tropft natürlich auch in die Kannenflüssigkeit, weshalb deren leichte Fluoreszenz logisch ist. Wenn die höhere Beutemenge unbehandelter Fallen mit solchen verglichen wird, deren Peristome mit Lösungsmitteln gewaschen oder schlicht abgeschnitten wurden, ist das als Beweis für die Fluoreszenz als Ursache recht untauglich. Logischerweise wurde da auch der Nektar entfernt und möglicherweise schädigte das Lösungsmittel sogar noch das Gewebe des Peristoms, was dessen Attraktivität zusätzlich heftig reduziert. Obwohl verschiedene Untersuchungen von unterschiedlich polaren Lösungsmitteleluaten der Peristome durchgeführt wurden, brachten diese außer Zucker (polare Glykoside oder phenolische Zuckerester) nur wenig Konkretes zutage. So werden z. B. komplexe Verbindungsklassen wie Chinone mittels allgemeiner Farbtests (mit KOH) nachgewiesen, bei denen es sich in Nepenthes bekannterweise um das für Mikroorganismen toxische Plumbagin (Derivat des Naphtochinons) oder die von Cannon et al. (1980) in den Wurzeln von N. rafflesiana isolierten und als Nepenthon A-E bezeichneten Naphtochinone handeln dürfte. Die werden jedoch alle gar nicht erwähnt. Die isolierte fluoreszierende Substanz, auf die es ja hier ankommt, wird lediglich als „Spuren phenolischer Verbindungen“ identifiziert, die in Methylenchlorid am Löslichsten waren. Mittels Dünnschichtchromatographie (HPTLC) wird der fluoreszierende Anteil zwar etwas genauer untersucht, jedoch leider ohne Vergleich mit den infrage kommenden Referenzsubstanzen. Leider wird so das eigentliche Ziel dieser Ursachenforschung verfehlt, nämlich die exakte Beantwortung der Fragen, welche optisch aktive Substanzen genau im Nektar enthalten sind, welche pharmakologische Wirkung deren Substanzklasse zugeschrieben wird, sowie natürlich ein Vergleich der in Sarracenia und Nepenthes (die nicht näher verwandt sind) für die Lumineszenz verantwortlichen Verbindungen. Was ist nun aber wirklich verantwortlich für die bläuliche Fluoreszenz des Nektars in unterschiedlichen Gattungen? Tatsächlich ist eine toxikologisch hochwirksame Substanz bei Sarracenia bereits seit Langem bekannt, sie wurde aber leider nicht einbezogen und gezielt untersucht. In der Tat haben Derivate heterozyklischer und polyzyklischer Aromaten auch vielfältige und starke pharmakologische Wirkungen bei Mensch und Tier, daher natürlich auch bei Arthropoden, der potenziellen Beute von Karnivoren. Wahrscheinlich handelt es sich also bei der optisch aktiven Substanz im Nepenthesnektar eher um eine Verbindung, deren toxikologische Wirkung für die Pflanze im Vordergrund steht. Die Fluoreszenz wäre dann lediglich ein verbreiteter Nebeneffekt solcher Substanzen, deren eigentlicher Zweck allerdings mit dem Entfernen der Fluoreszenz durch Lösungsmittel oder Abschneiden der Peristome auch wegfällt. Wie zuvor erwähnt, ist so ein stark bläulich fluoreszierendes Toxin schon sehr lange bekannt und wurde bereits in den Wurzeln von Sarracenia purpurea (Björklunt 1864) und im Nektar von Sarracenia flava (Mody et al. 1976) nachgewiesen. Es ist das für Menschen und Tiere neurotoxische Coniin, ein Piperidinderivat, besser bekannt als das Gift des Gefleckten Schierlings, das bereits Sokrates im alten Griechenland dahinraffte (Stichwort Schierlingsbecher). Coniin unterbricht die Übertragung der Nervensignale auf die Muskulatur, was zu Lähmungen führt, die sich beim Menschen in den Füßen beginnend nach oben im Körper ausbreiten. In der Medizin gelten Sehstörungen und Schwäche in den Füßen als typische Anfangssymptome einer oft tödlichen Schierlingsvergiftung. Bei Insekten bewirken bereits 100 ng (1 Nanogramm = 1 milliardstel Gramm) Coniin innerhalb von 30 Sekunden eine vollständige Lähmung (Juniper et al. 1989). Dadurch ist es eine perfekte Droge, um Beutetiere am Rand der Fallgruben durch Sehstörungen und Verlust der Beinkontrolle abstürzen zu lassen. Tatsächlich zeigen Fliegen und Wespen, die längere Zeit am Deckel von Sarracenia flava Nektar aufgenommen haben, ein auffällig träges und unkoordiniertes Verhalten. Zwar konnten wir in der einschlägigen Karnivorenliteratur keine Hinweise auf die Fluoreszenz des Coniins finden, in der chemischen Literatur wird das lange bekannte Nervengift jedoch als ausgesprochen intensiv bläulich fluoreszierend beschrieben und dürfte daher für die Lumineszenz des Nektars zumindest bei Sarrcenia verantwortlich sein. Der eindeutige Nachweis von Coniin oder ähnlicher Verbindungen (Piperidinalkaloide, zu denen auch das Nikotin gehört) in Nepenthes steht allerdings noch aus, weshalb dementsprechende Untersuchungen noch durchgeführt werden sollten. Da alle oben genannten und bereits bekannten toxikologisch wirksamen Substanzen in dem berühmten Buch The Carnivorous Plants (Juniper et al. 1989) aufgeführt sind, erscheint es etwas verwunderlich, dass diese in der Publikation von Kurup et al. gar nicht erwähnt werden, obwohl auch aus diesem Buch zitiert wird. Mit den zur Verfügung stehenden Analysemethoden wie z. B. der Massenspektroskopie (mit vorgeschaltetem Gaschromatographen) wäre ein einfacher Vergleich der Eluatspektren mit denen der bereits bekannten Wirkstoffe ja durchaus möglich gewesen. Immerhin kann man es als sehr positiv bezeichnen, dass die Publikation mit den Fotos der „leuchtenden Karnivoren“ in der Öffentlichkeit wie in Fachkreisen auf sehr großes Interesse stieß, weshalb dieser Beitrag entstand und zukünftig den leider unbeantwortet gebliebenen Fragen mit Sicherheit mehr Aufmerksamkeit zuteil wird. Für Sokrates war die Lumineszenz des tödlichen Gifts im Schierlingsbecher sicher völlig nebensächlich und bei Karnivoren, deren potenzielle Beute nach dem Genuss des vergifteten Nektars benebelt und mit Lähmungserscheinungen schneller in die Fallgruben plumpsen, ist das wahrscheinlich ähnlich.
Literatur Björklund, D. (1864). Untersuchungen aus dem Laboratorium der pharmaceutischen Gesellschaft in St. Petersburg. Sarracenia purpurea169, 93-124. (a) Chemische Untersuchung des Wurzelstockes. Archiv der Pharmacie (Hannover). Cannon, J.R., Lojanapiwatna, V., Raston, C.L., Sinchai, W. and White, A.H. (1980). The quinones of Nepenthes rafflesiana. The crystal structure of 2,5-dihydroxy-3,8-dimethoxy-7-methylnaphto-1,4-qinone (nepenthone-C). Australian Journal of Chemistry 33, 1073-1093. Juniper, B.E., Robins, R.J., and Joel, D.M. (1989). The Carnivorous Plants. Academic Press, London. Kurup, R., Johnson, A.J., Sankar, S., Hussain, A.A., Kumar, C.S., Baby, S. (2013). Fluorescent Prey Traps in Carnivorous Plants. Plant Biology Volume 15/2. Mody, N.V., Henson, R., Hedin, P.A., Kokpol, V. and Miles, D.H. (1976). Isolation of the insect paralyzing agent coniine from Sarracenia flava. Experimentia 32: 829-830. Warwicker, N. (2013). Carnivorous plant species glow blue to lure prey. BBC Nature News 19. Februar 2013. Prisma: Lockmittel Licht. Der Spiegel 10/2013 p. 121. |
||||||||||||||||||||||||||||||||||