published in Carnivorous Plant Newsletter (ICPS)
Vol. 39/4:101-113
(enhanced
with additional photos and video links on this website)
Introduction
Many CP-books describe the
trapping mechanism of Drosera generally as leaves
with sticky stalked
glands, which are able to bend towards the caught prey. However, the
traps are
actually not that simple. Even as early as 1875, Charles Darwin, in his
book
"The Insectivorous Plants" (Darwin
1888)
and a few later scientific publications mention two different tentacle
structures for Drosera rotundifolia. Since 2003, we
have published information
showing the morphology of Drosera tentacles is
actually even more
sophisticated (Hartmeyer 2003, 2004; Hartmeyer & Hartmeyer
2005, 2006a-d,
2008a, b). As of December 2009, we have investigated more than 100
world-wide
distributed species and varieties. From this work, we concluded that
the term
"stalked glands", often used as synonym for all Drosera
tentacles, must be considered as much too simplifying and even
incorrect in
many cases. A good example is the Australian Pimpernel Sundew (D.
glanduligera
Lehm.), published by Lehman in 1844. In the early 1970s, a
nine-year-old young
boy named Richard Davion found that the elongated marginal tentacles of
D.
glanduligera, which
was
growing close to his property in Adelaide, moved within fractions of a
second.
But like Lehman’s description, the carnivorous plant
literature (i.e. Erickson 1968 and
Lowrie 1989)
described the plant as a common sundew, and so for many years nobody
believed
this young Australian.
In 2003, Davion mailed his
private video-shots of the phenomenon, including some seeds, asking us
to
confirm his observation. Equipped with camcorder and USB-microscope, we
examined the minute, always hungry and absolutely nasty to cultivate
sundew,
during two winter seasons. By 2005, we had collected enough material to
fully
confirm Davion’s observations and to publish even more
amazing features, such
as the unique ontogenetic development (Hartmeyer & Hartmeyer
2005). We
were also able to
produce our Hunting Veggies® video and to describe
the conditions necessary to
cultivate that "diva", which is considered by some experts as a kind
of missing link between the sticky traps and the snap-traps in family
Droseraceae. Merging this awareness with our continuing experiments
with other
"snap-tentacle sundews" brought up suddenly, and to our surprise, a
clear view of the relations between, and even the phylogeny of, the
Droseraceae
based on their easily recognized different trap structures. One more
result was
the understanding of the morphology and function of the odd yellow
lens-tentacles on the leaves and bracts of D. hartmeyerorum
Schlauer
(Schlauer 2001). So now, almost four years after our DVD "Drosera:
Snap-Tentacles and Runway Lights" (Hartmeyer & Hartmeyer 2006)
was
shown at the ICPS conference 2006 in Frostburg - which documents this
work
until 2005 - , it’s a pleasure for us to now provide a
detailed summary of our
comparative examinations on Drosera tentacles for
CPN.
Drosera
tentacles in literature since 1875
Talking about the
ability of some species of
carnivorous plants to catch prey with active moving traps has been of
high
interest for a long time. Therefore, it is actually amazing that the
easily
observed fast movement, and the differing morphology, of marginal Drosera
tentacles, especially with several eye-catching pygmy Drosera
(section Bryastrum,
Lamprolepis), has been badly neglected, or simply ignored --
even with the
publication of new species in recent literature -- at least until our
publication in early 2005 on a sundew possessing a true snap-mechanism
(Hartmeyer & Hartmeyer 2005). The scientific book "The
Carnivorous
Plants" (Juniper et al. 1989)
mentions that several species exist which develop rapid moving,
glue-less
marginal tentacles (e.g.D.
burmannii), putting
them
into a context of evolution from Drosera tentacles
to the snap trap of
Dionaea. But as mentioned, even the old authors knew about
the
non-glandular tentacles of D. rotundifolia. Beside
Charles Darwin’s
first remarks in "Insectivorous Plants" (Darwin 1888),
descriptions are mostly restricted to a
few scientific publications in professional journals, such as Flora
(1904),
where C.A. Fenner (Germany) provided wonderful and very detailed
botanical
drawings of D. rotundifolia and other carnivorous
genera (Fenner 1904).
Investigations on the movement of Drosera tentacles
and how close they
are related to the trigger hairs of Aldrovanda and Dionaea
have
been published by Prof. Stephen Williams (USA) in the early 1970s
(Williams
1976, 2002; Williams & Pickard 1974, 1979). Using minute
electrodes, he
measured action potentials produced at the head or at the junction
between head
and stalk by touching or chemical stimulation (i.e.
with sodium chloride) of the tentacle head. The action
potentials move down the stalk where they cause particular cell-groups
to
increase their inner pressure. Without any junction, the whole tentacle
starts
bending inward toward the lamina due to rapid cell expansion (called
turgor-growth). Stephen Williams examined mucilage producing tentacles
of D.
capensis for his experiments. In the early 20th
century, Henry
D. Hooker, Jr. (1917) used the fitting term "non-glandular bisymmetric
tentacle head" and dedicated his research on "the mechanics of
movement in Drosera rotundifolia" mainly to the
bending stalks of
its marginal tentacles. In 1972, H.W.J. Ragetli, M. Weintraub, and
Esther Lo
(Ragetli et al. 1972)described
two tentacle structures for
the Drosera trap "… bisymmetrical
tentacles lying in the plane of
the leaf blade and extending outward from the leaf margin; and radially
or
nearly radially symmetrical tentacles extending upwards from the upper
surface
of the leaf … composed of an oval shaped head, a cylindrical
stalk, and a
connecting zone". However, it was worth it to examine numerous species
again, because our results show clearly that there are not only more,
but even
really amazing, adapted tentacles with clearly different morphology.
Drosera glanduligera, the snap-trap sundew
In 1994, we filmed Drosera
burmannii (section Thelocalyx) in our
greenhouse to record the rapid
movement of its non-glandular elongated marginal tentacles (Hartmeyer
&
Hartmeyer 2006). Surprisingly, we also found such tentacles with some
other
sundews standing nearby. Actually, less eye catching, but showing the
same
tactic, were the hybrids D. capensis x aliciae and D.
rotundifolia x spatulata
(synonym: D.
tokaiensis), some African species (section Drosera),
as well as some
pygmy Drosera (section Bryastrum/Lamprolepis). We found it
very interesting that some species showed this feature and others not.
We kept
the phenomenon in mind for a future project. In 2003, we received the
above mentioned parcel from Richard Davion. He wrote about "Ribbon
Tentacles" on D. glanduligera (monotypic section Coelophylla)
which need only fractional parts of a second to bend. In 1995 and 1999,
he even
published articles on the subject in "Flytrap News" (Davion 1995,
1999) mentioning also the movement of D. burmannii,
D. pygmaea,
and D. callistos. This
proved to be in vain, because obviously CP enthusiasts either ignored
or did
not believe him – until we showed his VHS recordings on our
DVD "A Hunting
Veggies Cocktail" (Hartmeyer & Hartmeyer 2004). For our own
experiments, we used a PAL video camera (25 pictures per second) for
speed
measurements and were able to prove with single pictures that,
triggered with a
tooth pick or tweezers, D. glanduligera moves its
elongated marginal
tentacles within sensational 0.16 (+/- 0.04) seconds like a snap trap,
which
set a unique record for the genus and is as fast as the Venus Flytrap
snaps
shut. In addition, our attendant microscope pictures show a clearly
distinct
tentacle morphology (see below), again unique for the whole genus.
Also, the
ontogenetic development of its seedlings is clearly different from
other
species. So we thoroughly confirmed Davion’s discovery, and
wrote an article
for the German CPS (GFP) newsletter Das
Taublatt (Hartmeyer & Hartmeyer 2005). With our
publication, we
coined the term "snap-tentacle" in English and, after a proposal by
Dr. Jan Schlauer, the term "Schnelltentakel" in the German language.
A new video showing the snap-tentacles of D. glanduligera
in action can
meanwhile be seen on YouTube:
A new diversity of tentacles
Already in the early state
of our experiments with rapid moving marginal tentacles, but
increasingly after
our examination of D. glanduligera, our macro-shots
and microscope
pictures disclosed a clandestine diversity, demanding an even closer
inspection. Amazed about different shapes, speed, and the spasmodic
appearance
even in closely related species, we realized that these results contain
not
only obvious information about adaptation and relationship within the
genus,
but also on the evolution of the whole family Droseraceae.
Figure A1 (additional): Drosera hartmeyerorum lens-tentacles on florescence and leaf base. Video-screenshot by I.&S.Hartmeyer.
Figure A2 (additional): Drosera hartmeyerorum lens-tentacles illuminated with a torch. Video-screenshot by I.&S.Hartmeyer.
Figure A3 (additional): Drosera hartmeyerorum lens-tentacles on contrasting red bract. Video-screenshot by I.&S.Hartmeyer.
Figure A4 (additional): Drosera hartmeyerorum lens-tentacles magnified. SEM picture by Dr. Regina Kettering.
Beside the
"normal" symmetrical mucilage-producing tentacles growing upright on
the lamina, we identified three different types of marginal tentacles.
We also
found additional non-glandular structures like the unique
yellow-shining
lens-headed tentacles at the base of the trapping leaves and at the
bracts of
the florescence of D. hartmeyerorum (section Arachnopus).
The
head consists of lens-like transparent giant cells focusing at a bright
yellow
cell-structure in the center. Reflecting the incoming sunlight, these
lens-tentacles function like a yellow rear reflector (Hartmeyer
& Hartmeyer
2006). Additional lens-tentacles, placed at intervals on the dark red
bracts of
the green florescence, look like a chain of runway lights from above,
attracting insects (we found lots of baby grasshoppers) to walk or skip
down
the flower stalk into the dark red and sticky plant center from where
bundles
of lens-tentacles show the yellow lightning with high contrast. We also
found
different small 0.1 to 1 mm-sized emergences with a simple yellow
hemisphere-shaped head on the trapping leaves and stalk of Australian D.
indica, and translucent mushroom-like emergences with a waved
flat head on
a large African D. indica (Hartmeyer 2003;
Hartmeyer & Hartmeyer
2006). We guess that the
mushroom-like structures produce the sweet odor of that intensely
scented
plant. From personal communication, we know about some more different
emergences on sundewswhich are still subject to
further examination and
have not been published yet.
Our examinations on the lens-tentacles are on YouTube:
These findings produce an
increased understanding about the connection of marginal tentacles with
the
other genera assigned to the Droseraceae: Aldrovanda
and Dionaea.
Their modified non-glandular tentacles should not be forgotten. These
are the
trigger hairs on the snap-traps of both genera that developed from Drosera
tentacles, as Stephen Williams described in the 1970s. But, also, the
spine-like teeth on the margin of Dionaea lobes are
obviously a remnant
of marginal tentacles which lost their (no longer needed) trigger
function.
Three different
types of marginal
tentacles
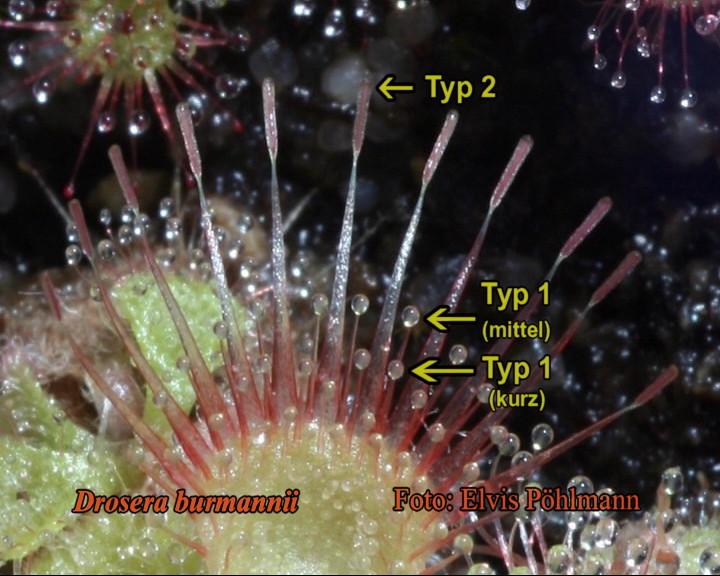
Figure 1: Drosera scorpioides,
T1 sticky marginal
tentacles. Photo by E. Pöhlmann.
Concerning the rapid
moving
marginal tentacles of Drosera, we confine ourselves
only to the three
types of "modern" tentacles that all emerge with a very broad and
powerful base (four to six times broader than the base of the
cylindrical
tentacle stalks on the lamina) lying in the plane of the leaf blade.
The
following definition was published in 2008 (Hartmeyer &
Hartmeyer 2008;
McPherson 2008).
The
first type
of marginal
tentacles (T1, present mainly
on
the leaves of erect growing species) possess a mucilage producing,
glandular
symmetrical head which is similar to the normal stalked mucus secreting
glands
on the lamina of that species, however, develops on an elongated stalk
that emerges
with a very broad base. This type of elongated marginal tentacle is
typified on
the lamina of D. scorpioides for example. Movement
in this type of
tentacle takes place by turgor growth.
The
second type
of marginal
tentacle (T2, present mainly
on
the leaves of basal rosettes) possess a bisymmetrical, non-glandular
head that
lacks a sticky mucus secretion, often looking somewhat like a shovel
with a
flat lower side and an elevated mostly red, rarely green colored
cushion-like
glandless cell structure on the upper side, which develops on an
elongated
stalk that emerges with a very broad base. The size and shape of the
elongated
head and stalk vary considerably within different species. This type of
elongated marginal tentacle is typified on the lamina of D.
burmannii
for example. Important: T2- tentacles always lack secretions of glue
and
movement takes place by turgor growth.
Figure 2: Drosera
burmannii, T2 + T1 tentacle
pattern on adult plant. Annotated by S. Hartmeyer.
Figure 3: Drosera
glanduligera, T3 + T1
tentacle pattern on adult plant. Annotated by S. Hartmeyer.
The
third type
of marginal
tentacle (T3) is known only
from D. glanduligera. The stalk consists of a broad
lower section emerging
with a very broad base from the margin of the lamina and, connected by
a kind
of junction, a very slender but longer upper portion that terminates in
a
bisymmetrical, non-glandular head that lacks sticky mucus. Evidently,
the
tentacle head contains a trigger mechanism, which when activated,
causes the
tentacle to move very rapidly. The slender upper part of the tentacle
bends
forward from the junction-like midsection so that the head of the
tentacle
snaps towards the middle of the lamina. The whole process takes place
remarkably quickly; we measured 0.16
seconds. The process
of movement and the morphology of T3-tentacles differ therefore from
all other
tentacles known from the foliage of species of Drosera.
The mechanism by
which movement takes place is not currently understood. It is possible
that
some turgor growth takes place in order for the tentacle to bend,
however, the
process overall is much more sophisticated and therefore D.
glanduligera
is (until today) the only sundew that moves with the same speed as the
Venus
Flytrap.
We define a snap-tentacle
(T2 + T3) as a structure which emerges with a powerful broad base from
the
margin of the lamina, consists of an elongated stalk with or without
junction,
and a bisymmetrical non-glandular head. This definition includes the
second and
third type of elongated marginal tentacles described above, but not the
first
type, as despite its powerful broad base and often rapid movement, T1
carries a
mucilage producing, glandular symmetrical head (much like those of
regular
tentacles common to the leaves of all species of Drosera).
Any tentacle
with a conventional mucus secreting head is therefore not a
snap-tentacle.
Both
types of
snap-tentacles are able to bend rapidly, although through different
processes
and at very different speeds. T3-tentacles move 50 to 100 times faster
than
T2-tentacles. Since both types are non-glandular, it seems likely that
they
primarily play the role of ensuring and fixing prey to the center of
the lamina,
and so, to complicate an escape and/or to avoid the theft of prey by
clepto-parasites. Not to forget, usually behind the front row of
snap-tentacles
follow 1-3 rows with smaller and more upright-held sticky T1 tentacles,
which
develop also with the typical broad base (unlike the normal upright
glue-tentacles on the lamina) and support the snap-tentacles in fixing
prey.
Figure 5: Drosera
glanduligera, outstretched
snap-tentacle with junction in the middle.
Photo by S. Hartmeyer.
Figure 6: Drosera
glanduligera, triggered
snap-tentacle, bend at the junction.
Photo by S. Hartmeyer.
Table 1. Distribution of snap-tentacles (T2
& T3) in Drosera
sections (incl. seedlings)
Section
Snap-tentacle type
Section
Snap-tentacle type
Arachnopus
- - -
Meristocaulis
T2
Arcturi
- - -
Phycopsis
T2
Bryastrum/Lamprolepis
T2
Prolifera
- - -
Coelophylla
T3
Ptycnostigma
T2
Drosera
T2
Regia
- - -
Ergaleium
- - -
Stelogyne
T2
Lasiocephala
T2
Thelocalyx
T2
Archaic and modern Drosera
During our experiments, it
became clear that the ontogenetic development of the seedlings also
deserves a closer
investigation. Surprisingly, many seedlings possess leaf generations
(which
follow the cotyledons) with a minimum of three to five powerful
snap-tentacles
(T2) which emerge morphologically as outgrowth of the lamina, but many
of the
adult plants do not.
Unfortunately, we were not
able to get seeds from D. arcturi, D.
regia, and section Prolifera.Therefore, these species
were examined for
snap-tentacles looking at the first leaves of small plantlets, which
grew from
root- or leaf-cuttings (D. adelae, D. regia, and D. schizandra),
developed at the florescence (D. prolifera), or
opened first from winter
resting buds (D. arcturi). The results show clearly
that juvenile plants
of the oldest known species in the genus, D. arcturi
and D. regia,
never develop broad-based marginal tentacles (T1-T3). This confirms the
assumption that the first sundews caught their prey only with simple
glue-tentacles on the lamina, but had already developed the ability to
roll the
complete leaf.
Also, the species in section Arachnopus
(D. hartmeyerorum, D. indica),
and section Prolifera
(D. adelae, D. prolifera, D.
schizandra) seem to descend
from archaic species because here we found no snap-tentacles. However,
one
exception in section Prolifera is D.
adelae, which
possess some tentacles at the
leaf-tips which also emerge from a broad base and are able to move
faster (45 –
90 seconds) than the other tentacles on the lamina; therefore, they
show some
similarity to T1 tentacles. This may be a sign that section Prolifera
is
somehow in between the archaic and modern Drosera,
but its tentacle
structure is more closely related to the modern section Drosera
than to
section Arachnopus. This would also subsequently
confirm Jan Schlauer's
decision (Schlauer 1996) to extract the Queensland sundews from section
Arachnopus
into its own section Prolifera. Unlike all species
which developed
elongated marginal tentacles to fix prey to avoid theft by
clepto-parasites,
the species in section Arachnopus adapted and
optimized their tentacles
obviously to increase the attraction of prey.
The tuberous sundews
(section Ergaleium) deserve further attention.
Neither seedlings nor
adult plants possess snap-tentacles (T2 + T3), but mainly the erect
species
show typical broad-based marginal T1-tentacles. As we documented with D.
macrantha (Hartmeyer & Hartmeyer 2006), when
triggered, they bend with
about the speed of D. burmannii. This is actually a
very modern feature.
Without a doubt, something
substantial must have happened in the past that triggered the
development of
powerful broad-based marginal tentacles to fix prey to the lamina,
maybe to
improve digestion, but more likely to complicate the theft of prey.
With this
important adaptation, the modern Drosera turned up,
becoming so
successful that nowadays all (examined) species world-wide assigned to
the
sections Bryastrum/Lamprolepis, Drosera,
Lasiocephala, Meristocaulis,
Phycopsis, Ptycnostigma,
Stelogyne,
and Thelocalyx descend from
the first modern sundews, all starting their life as a seedling
equipped with
powerful T2 snap-tentacles.
Therefore, we define all
species which develop either T1- or T2- or T3-tentacles as modern Drosera
and those without these broad based marginal tentacles as archaic Drosera
(see
Table 1).
As mentioned above, D.
glanduligera develops very effective and unique tentacles. At
a first
glance, the fastest snap-mechanism of the genus seems to be the most
sophisticated and, therefore, very modern. Using a magnifying glass, we
observed that minute springtails were catapulted into the sticky center
of the
lamina after touching the unique snap-tentacle extending from the
lamina --
like the alarm threads of some spiders. But strange enough, its
seedlings start
with first leaf-generations (after the cotyledons) that possess only T1
stalked
glands, even the next two or three leaf generations show only gluey and
intermediate (between T1 & T3) tentacles. Only from the fourth
or fifth
leaf generation on, the typical rapid moving snap-tentacles with the
junction-like middle-part function are found. Therefore, the
ontogenetic
development is distinct from all other species (examined so far) in the
genus
and therefore D. glanduligera must be considered as
the oldest of the
described modern sundews.
Basal rosettes and erect plants
In section Bryastrum/Lamprolepis, for example, we find a lot of adult
plants that possess very prominent snap-tentacles, but closely related
species
growing sometimes at the same site, do not. This confusion disappears
immediately if the growing shape is taken in consideration, as shown in Table 2. This pattern
is typical for the species
assigned to the modern sections and shows clearly that snap-tentacles
are very
important for seedlings and basal rosettes, but obviously not for erect
plants.
If the fixing of prey to the lamina would have been developed only to
improve
digestion, it should be useful for both basal and erect plants, but if
the
fixation is a strategy to avoid the theft of prey - which happens
frequently,
for example, by ants - the strategy is only important for trapping
leaves near
the soil (i.e., seedlings and basal
rosettes). Erect growing plants, like D. scorpioides
or D.
madagascariensis,
but also
erect-held leaves, such as on adult D. capensis or D.
binata (all
seedlings and juvenile plants possess T2-tentacles), catch mostly
flying
insects, for which their sticky T1-tentacles work much more
effectively. Just a
little contact with the tip of a wing is sufficient to paste and hold a
mosquito. The speed of movement achieved by T2-tentacles may be
impressive for
a plant, but is certainly not dangerous for flying insects.
Drosera of the Northern
Hemisphere
Figure A5 (additional): Drosera rotundifolia bisymmetrical tentacle head. USB-microscope picture by S. Hartmeyer
Figure A6 (additional): Drosera intermedia hibernacula. Video scrennshot by S. Hartmeyer
Referring to the whole
genus, only a small number of species grow in the Northern Hemisphere.
While
species like D. capillaris and D.
spatulata remain relatively
south, preferring warmer temperatures, D. anglica, D.
intermedia, D. linearis, D.
rotundifolia,anda
little less tough D. filiformis,
show a perfect adaptation to frost and short growing seasons by
developing a
true dormant bud. All species are assigned to the modern section Drosera
and all of their seedlings possess snap-tentacles. However, the only
typical
basal rosette is D. rotundifolia, and confirming
the above mentioned
pattern for the growing shape, only this true northern species develops
T2-tentacles on adult leaves -- not always and not every leaf, which
caused a
little confusion when we examined our greenhouse plants for our video
and found
not one snap-tentacle on any of our plants. But in the next season -
after the video
has been edited - the same plants developed them. None of the species
growing
in colder regions of the Southern Hemisphere, neither the simple winter
bud of D.
arcturi nor the thickened roots of D. binata
or D. cistiflora
are able to survive temperatures below -20 or even -30 degrees
centigrade,
which is no problem for the specialized hibernacula of the northern
species. A
less effective adaptation to frost in the Southern Hemisphere is not
illogical
if one considers that the northern ice shields moved much further south
(and
back), occupying much more land than the Antarctic ice shields, which
show even
today still about 80% of their maximum extension during the last
glacial period
about 20,000 years ago. So we can assume that the species of the
Northern
Hemisphere developed their strategies to survive extreme frost during
the
approximately 30 glacial periods within the last 2 million years.
Drosera of the Southern
Hemisphere
The predominate number
of Drosera
can be found in the Southern Hemisphere. D. regia
in South Africa and D.
arcturi in Australia/New Zealand are the oldest known species
and most
experts agree that the genus developed in the south, spreading north
over the
course of time. As mentioned above, only modern species can be found
north of
Australia and South Africa, and surprisingly, the shape of their
T2-tentacles
seems to provide further information about their history. However, most
T2-heads on seedlings and young leaves are round, or in D.
binata
sometimes nearly square, so only the shape of heads on adult leaves
should be
compared.
Let’s start in Australia,
where we find more than 100 of the approximately 180 species world-wide
(estimated, because new species are frequently found). As mentioned
above, D.
glanduligera , due to its ontogenetic development, is
probably the oldest
of the modern Drosera. The greatest morphological
similarity with the
unique T3-tentacles of D. glanduligera, and
therefore indicating the
closest relationship, are the T2-tentacle heads of pygmy Drosera,
where
we find also very long slender stalks with minute round heads.
Actually, we
also found the fastest T2-movement in this section with D.
ericksoniae.
Five seconds to bend to the lamina is indeed fast, however, still a
magnitude
slower than D. glanduligera. The large dichotomous
leaves of D.
binata (section Phycopsis) possess only
mucus-producing
T1-tentacles, but seedlings and juvenile plants develop T2-tentacles
until the
lamina starts to divide.
Drosera
burmannii, with very
prominent snap-tentacles, shows the largest rectangular heads in the
genus,
referring to the plant size. It was in the past often called the
fastest moving
sundew, but with 8-15 seconds measured it is not faster than many
pygmys. The
only species in Australia assigned to section Drosera
showing
rectangular heads is D. spatulata. But both D.
burmannii and D.
spatulata also grow further north in Asia and seem to be not
typical of
Australians. The clear majority there show round to slightly oval heads.
Most of the species growing
in the Americas are assigned to section Drosera.
Unfortunately, we had
only few plants to examine, but it's interesting that several species
like D.
montana, D. capillaris, D. felix, but
also young plants of D.
ascendens and D. villosa (the last two
species have only
T1-tentacles when matured) show a somewhat more three-cornered than
true
rectangular shape, which seems to be an American feature. But, as in
Australia,
we find also a few species that do not fit. Drosera
sessilifolia
(section Thelocalix) is only hard to distinguish
from D. burmannii
in Australasia and its snap-tentacles move with the same speed. The
recently
discovered D. meristocaulis (section Meristocaulis)
develops
snap-tentacles and is surprisingly very similar to the Australian pygmy
Drosera. Drosera rotundifolia
growing only in colder climates from North America
through Europe and Siberia to Japan shows more rectangular heads.
Figure A7 (additional): Drosera sessilifolia adult trap with snap-tentacles. Video-screenshot by I.&S.Hartmeyer
Most of the African species
are also assigned to section Drosera, but D. admirabilis,
D. aliciae, D. cuneifolia,
and most of the other examined plants possess large rectangular
T2-tentacle
heads like D. rotundifolia or D. spatulata.
In this connection
also, the rectangular heads of D. burmannii
(Australasia) and D.
sessilifolia (South America) should be mentioned again. A
good example for
the development of modern tentacles during the maturation of erect
plants is
the South African D. cistiflora (section Ptycnostigma)
which has
first round, later drop-shaped T2-heads as long as it grows as basal
rosette,
but only two or three leaf-generations after the erect growth started,
only
sticky T1-tentacles appear in their place.
Looking at the morphology,
the majority of species in Australia develop round-shaped T2-heads; in
Africa,
we find mainly rectangular heads; and in the Americas, the lower part
of the
cushion-like cellstructure is often a little smaller than the upper
part, so
they appear more wedge-shaped. That looks very much like a separated
development on each continent, but from common ancestors. A
development, that
is similar and well known also from other plant genera (i.e.Drosera,
we have
- even without usable fossils - a good evidence that carnivory is
actually a
very old strategy within the plant kingdom, existing even in Gondwana,
the
giant continent in the Southern Hemisphere which broke apart into
Antarctica,
Australia, Africa, India, and South America between 130 - 100 million
years
ago, so the archaic sundews must even be considerable older. This
theory would
also provide a logical explanation for the occurrence of obviously
closely
related species on different
(now) remote continents (i.e. Proteaceae).
Table 2. Examples for tentacle types in relation with
the
growing shape
Section
Species
Seedling
Adult
Growing shape
Arachnopus
D. hartmeyerorum
- - -
Lens-tentacles
Erect
"
D. indica (all
varieties, probably even undescribed species)
- - -
different small emergences
Erect
Arcturi
D. arcturi
- - -
- - -
Rosette, erect leaves
Bryastrum/Lamprolepis
D. callistos
T2
T2
Basal rosette
"
D. ericksoniae
T2
T2
Basal rosette
"
D. occidentalis
T2
T2
Basal rosette
"
D. pulchella
T2
T2
Basal rosette
"
D. pygmaea
T2
T2
Basal rosette
"
D. dichrosepala
T2
T1
Erect
"
D. enodes
T2
T1
Erect
"
D. lasiantha
T2
T1
Erect
"
D. scorpioides
T2
T1
Erect
Coelophylla
D. glanduligera
T1
T3
Basal rosette
Drosera
D. aliciae
T2
T2
Basal rosette
"
D. capensis
T2
T1
Erect
"
D. madagascariensis
T2
T1
Erect
"
D. ramentacaea
T2
T1
Erect
"
D. rotundifolia
T2
T2
Basal rosette
"
D. spatulata
T2
T2
Basal rosette
Ergaleium
D. erythrorhiza
- - -
- - -
Basal rosette
"
D. bulbosa
- - -
- - -
Basal rosette
"
D. lowriei
- - -
T1
Erect
"
D. macrantha
- - -
T1
Erect
"
D. modesta
- - -
T1
Erect
"
D. peltata
- - -
T1
Erect
Lasiocephala
D. lanata
T2
T1
Rosette, erect leaves
"
D. ordensis
T2
T1
Rosette, erect leaves
Phycopsis
D. binata
T2
T1
Rosette, erect leaves
Prolifera
D. adelae
- - -
around leaf apex resemblance with T1
Basal rosette
"
D. prolifera
- - -
- - -
Rosette, erect leaves
"
D. schizandra
- - -
- - -
Basal rosette
Ptycnostigma
D. cistiflora
T2
T1
Erect
Regia
D. regia
- - -
- - -
Rosette, erect leaves
Thelocalyx
D. burmannii
T2
T2
Basal rosette
"
D. sessilifolia
T2
T2
Basal rosette
Conclusion
If we summarize all of the
observations mentioned above and combine them
with scientific confirmed incidents, a surprising theory (to be
confirmed by
other scientific disciplines) can be expounded:
Once upon a time, 200 or maybe
even 300 million years ago, archaic vertebrates, plants, and arthropods
like
millipedes, insects, mites, and spiders, were living in the tropical
rainforests at the southern coastal regions of Gondwana, an area that
corresponds today mainly with Antarctica, but also the southern tip of
South
America in the west and Australia in the east. Certainly different from
today,
but following the same principles of life as nowadays, food chains and
a web of
interdependence including mutualism and parasitism developed. This
interrelation enabled a group of early angiosperms (flowering plants)
to use
the manifold arthropods for their sexual reproduction. Compared with
pollination by wind, animal-assisted pollination is much more
beneficial
because only a fraction of the pollen is necessary if it is carried by
animals
specific to the females. As a result, the modern insect-attracting
flowers and
a new web of interdependences developed.
Today all known carnivorous
plants are angiosperms, so it is only logical to consider that they
also
realized very soon that arthropods attracted by provided nectar are not
only
good to transport pollen, but are also compact protein-packs which can
be used
very efficiently as nutrients, providing a certain advantage on poor
rainforest
soils. Therefore, some angiosperms started to catch prey with sticky
tentacles
on the surface of their leaves (and others probably also with pitcher
forming
leaves) and learned in the course of time to produce digesting enzymes
and
glands to absorb the nutrients. Thus, the archaic sundews, looking
probably
somewhat like D. schizandra or D. regia
started their successful
existence.
About 130-140 million years
ago, these archaic Drosera had conquered the whole
south of Gondwana,
but now some wasps decided to settle on the ground and to use wings
only for
their mating flights. As a result, one of the most successful genera on
earth
appeared: the ants. Without a doubt, the appearance of ants caused a
kind of
revolution for all organisms living on the ground. Even today, it can
be
frequently observed that ants, which are certainly also prey for Drosera,
steal considerable amounts of prey that the sticky plants caught. So,
with the
ants, the quantity of nutrients became suddenly seriously reduced. The
ability
to roll in leaves to hold captured prey more effectively and to
increase the
surface for digestion was too slow to avoid the theft by the
omnipresent ants
(and/or other clepto-parasites). But as part of the never ending story
in
nature to become the fittest survivor, some Drosera
developed marginal
tentacles equipped with a powerful broad base and the ability to move
rapidly
enough to fix the prey effectively before predatory ants are able to
steal it.
With this successful adaptation, the modern Drosera saw
the light of
day. During the following millions of years, new Drosera
species and
even two new genera changed the sticky and snap-tentacle armed trap
into a
closing snap-trap (which is even more effective to prevent the theft of
prey),
prospered well and proliferated north.
Caused by the continental
shift, Gondwana broke asunder into the new southern continents and as
early as
100 million years ago, the oceans in between became so large that the
now
separated species on the divided landmasses developed independent from
each
other. Antarctica, the area where probably the first Drosera
developed,
became isolated in the polar region and the archaic life was killed by
frost.
Australia, the last continent that broke away from the east of
Antarctica
drifted toward the equator and carried so many Gondwana species, like
the
marsupials, that it is now the epicenter with the most descendants of
archaic
and modern Drosera. South America broke away from
the west of Antarctica
earlier, but different species like D. sessilifolia
and D.
meristocaulis, with
obviously closely related plants in Australia, may be evidence for
common
ancestors in Gondwana. Also, in South Africa, which separated even
earlier, D.
indica and numerous modern species survived, including the
most archaic D.
regia, which has several similarities with the Australian
(and New Zealand) D. arcturi.
When our first observations
and investigations on the movement and different shapes of elongated
marginal
tentacles started, it was interesting from the beginning, but we never
expected
how much new information can be collected even today using a simple
microscope.
Most of our published results have meanwhile been confirmed and
supplemented by
other CP-experts and authors. However, as mentioned before, our
conclusion
supporting the so called "Gondwana theory" should soon be confirmed
by other scientific disciplines.
Acknowledgements
The following persons,
listed in alphabetical order, provided helpful comments or other
assistance in
the preparation of this paper: Greg Bourke, Richard Davion, Christian
Dietz,
Andreas Fleischmann, Anja & Holger Hennern, Regina Kettering,
Elvis
Pöhlmann, Barry Rice, Jan Schlauer, Stephen Williams, and some
more anonymous
contributors.
References
Darwin, C. 1888. Insectivorous
Plants. 2nd edition, revised by Francis Darwin.
John Murray, London.
Davion, R. 1995. Now you see it
-
Now you don’t. Flytrap News (CPSNSW) 8/4: 17.
Davion, R. 1999. That damned
elusive
Pimpernel. Flytrap News (CPSNSW) 13/1: 10.
Erickson, R. 1968. Plants of
Prey.
University of Western Australia Press, Nedlands.
Fenner, C.A. 1904. Beiträge zur Kenntnis
der
Anatomie, Entwicklungsgeschichte und Biologie der Laubblätter
und Drüsen
einiger Insektivoren.
Flora
1904, 93 Band.
Hartmeyer, S. 2003. Neues
vom Sonnentau: Die Sektion Arachnopus
unter dem Mikroskop. Das Taublatt (GFP)
2003/3:
4-8
Hartmeyer, S. 2004. Drosera
section Arachnopus under the
microscope. Carniflora Australis (AUSCPS)
2004/3: 11-17.
(modified English translation,
published in Australia).
Hartmeyer, I., and Hartmeyer,
S.
2004. Quer durch’s
Karnivorenbeet. (English:
A Hunting Veggies
Cocktail), DVD
Hartmeyer, I., and
Hartmeyer, S. 2005. Drosera glanduligera:
Der Sonnentau mit "Schnapp-Tentakeln". Das Taublatt (GFP) 2005/2:
34-38.
Hartmeyer, I. and Hartmeyer, S. 2006a. Verborgene Vielfalt: Die
Schnelltentakel der Gattung
Drosera.Das Taublatt
(GFP) 2006/1: 38-50.
Hartmeyer, I., and Hartmeyer,
S. 2006b.
Clandestine diversity: Snap-tentacles of genus Drosera.
Carniflora
Australis (AUSCPS)
2006/No 7:4-18.
(modified English
translation, published in Australia).
Hartmeyer, I., and Hartmeyer,
S. 2006c. La
diversità nascosta: i
tentacoli a scatto nel genere Drosera - Traduzione
italiana di Chiara
dB.
AIPC Magazine (AIPC)
2006/3:14-25. (Italian
translation by Chiara dB,
published in Italy).
Hartmeyer, I., and Hartmeyer,
S. 2006d. Drosera –
Snap-Tentacles and Runway-Lights (German language: Drosera
-
Schnelltentakel und Landescheinwerfer), DVD.
Hartmeyer, I., and Hartmeyer,
S.
2006e. Drosera
hartmeyerorum
– Der Sonnentau mit Lichtreflektoren. Das
Taublatt (GFP) 2006/3: 4-9.
Hartmeyer, I., and Hartmeyer,
S. 2008a. Urtümliche und moderne
Tentakel unter dem Mikroskop. Das Taublatt (GFP)
2008/3: 36-43.
Hartmeyer, I., and Hartmeyer,
S. 2008b. Triple E meets Triphyophyllum
(German language: Triple E trifft Triphyophyllum),
DVD.
Hooker, Jr., H.D. 1917.
Mechanics of movement in Drosera rotundifolia.
Bul. Torrey Bot. Club 44: 389-403.
Juniper, B.E., Robins, R.J.,
and Joel, D.M. 1989. The Carnivorous
Plants. Academic Press, London: 303.
Lowrie, A. 1989. Carnivorous
Plants of Australia. Vol. 2 and 3.
University of Western Australia Press, Nedlands.
McPherson, S. 2008. Glistening
Carnivores - The Sticky-Leaved Insect-Eating Plants. Redfern Natural
History
Productions. Poole, Dorset, England
Ragetli, H.W.J., Weintraub, M.,
and Ester Lo. 1972. Characteristics of Drosera
tentacles. I. Anatomical and cytological detail.
Can. J. Bot. 50:
159-168.
Schlauer, J. 1996. A
dichotomous key to the genus Drosera L.
(Droseraceae), Carniv. Pl. Newslett. 25: 67-88.
Schlauer,
J. 2001. Drosera
hartmeyerorum spec. nov (Droseraceae), a new sundew in
sect. Arachnopus from Northern Australia.
Carniv.
Pl. Newslett. 30:
104-106.
Williams, S.E. 1976.
Comparative sensory physiology of the Droseraceae -
the evolution of a plant sensory system.
Proc. Amer. Philos. Soc.
120(3):
187-204.
Williams, S.E. 2002. Comparative
physiology of the Droseraceae sensu
stricto - How do tentacles bend and traps close?
Proc. 4th Intl.
Carniv. Pl. Conf., Tokyo: 77-81.
Williams, S.E., and Pickard,
B.G. 1974. Connections and barriers between
cells of Drosera tentacles in relation to their
electrophysiology.
Planta (Berl.) 116: 1-16.
Williams, S.E., and Pickard,
B.G. 1979. The role of action potentials in
the control of capture movements of Drosera and Dionaea.
Plant
Growth Substances. Springer Verlag, Berlin-Heidelberg-New
York.